
|

|
Большая Советская Энциклопедия (цитаты)
|

|
|
|
|
Ферменты | Ферменты (далее Ф) (от лат. fermentum – закваска), энзимы, специфические катализаторы, присутствующие во всех живых клетках. Почти все биохимические реакции, протекающие в любом организме и в своем закономерном сочетании составляющие его обмен веществ, катализируются соответствующими Ф Направляя и регулируя обмен веществ, Ф играют важнейшую роль во всех процессах жизнедеятельности.
Как всякие катализаторы, Ф снижают энергию активации, необходимую для осуществления той или иной реакции, направляя ее обходным путем – через промежуточные реакции, которые требуют значительно меньшей энергии активации. Так, реакция АБ ® А + Б в присутствии Ф идет следующим образом: АБ + Ф ® АБФ и далее АБФ ® БФ + А и БФ ® Б + Ф Например, для осуществления реакции гидролиза дисахарида в результате которого образуются и фруктоза, без участия катализатора требуется 32 000 кал (1 кал = 4,19 дж) на моль Если же реакция катализируется Ф b-фруктофуранозидазой, то необходимая энергия активации составляет всего 9400 кал. Подобное понижение энергии активации под влиянием Ф – следствие перераспределения электронных плотностей и некоторой деформации молекул субстрата, происходящей при образовании промежуточного соединения – фермент-субстратного комплекса (АБФ). Эта деформация, ослабляя внутримолекулярные связи, приводит к понижению необходимой энергии активации и, следовательно, ускоряет течение реакции (см. Катализ, Ферментативный катализ).
История изучения ферментов. В 1814 рус. химик К. Г. С. Кирхгоф открыл ферментативное действие водных вытяжек из проросшего ячменя, расщеплявших крахмал до Можно считать, что эти работы положили начало энзимологии (ферментологии) как самостоятельному разделу биологической химии. В 1833 французскими химиками А. Пайеном и Ж. Персо впервые был выделен из солода препарат фермента амилазы, что способствовало развитию препаративной химии Ф В середины 19 в. разгорелась дискуссия о природе брожения между Л. Пастером, с одной стороны, и Ю. Либихом, П. Э. М. Бертло и К. Бернаром – с другой. Опираясь на свои классические работы, Пастер развивал представление о том, что брожение вызывается лишь живыми микроорганизмами и что процесс брожения неразрывно связан с их жизнедеятельностью. Либих и его сторонники, отстаивая природу брожения, считали, что оно является следствием образования в клетках микроорганизмов растворимых Ф, подобных выделяемой из солода амилазе. Однако все попытки выделить из разрушенных дрожжевых клеток растворимый Ф, способный вызвать брожение, не удавались. Дискуссия Либиха и Пастера о природе брожения была разрешена в 1897 Э. Бухнером, который, растирая дрожжи с инфузорной землей, выделил из них бесклеточный растворимый ферментный препарат (названный им зимазой), вызывавший спиртовое брожение. Открытие Бухнера утвердило материалистическое понимание природы брожений и имело большое значение для дальнейшего развития как энзимологии, так и всей биохимии.
В начале 20 в. Р. Вильштеттер с сотрудниками стал широко применять для выделения и очистки Ф метод адсорбции (впервые предложен А. Я. Данилевским для разделения Ф поджелудочной Работы Вильштеттера, имевшие большое значение для характеристики свойств отдельных Ф, привели вместе с тем к принципиально неправильному выводу, что Ф не принадлежат ни к одному из известных классов органических соединений. Выдающимся успехом в выяснении природы Ф были исследования американских биохимиков Дж. Самнера, выделившего в 1926 в виде Ф уреазу из семян канавалии, и Дж. Нортропа, получившего в 1930 протеолитического Ф пепсина. Работы Самнера и Нортропа указали путь получения высокоочищенных препаратов Ф и вместе с тем неопровержимо доказали природу Ф
С середине 20 в. благодаря развитию методов физико- анализа (главным образом хроматографии) и методов химии расшифрована первичная структура многих Ф Так, работами американских биохимиков С. Мура, У. Стайна и К. Анфинсена показано, что Ф рибонуклеаза из поджелудочной быка представляет собой полипептидную цепочку, состоящую из 124 аминокислотных остатков, соединенных в 4 местах дисульфидными связями.
С помощью рентгеноструктурного анализа расшифрована вторичная и третичная структура ряда Ф Так, методом рентгеноструктурного анализа английский ученый Д. Филлипс в 1965 установил трехмерную структуру Ф лизоцима. Показано, что многие Ф обладают также четвертичной структурой, т. е. их молекула состоит из нескольких идентичных или различных по составу и структуре субъединиц (см. Биополимеры).
Общая характеристика ферментов. Все Ф разделяются на две большие группы: однокомпонентные, состоящие исключительно из и двухкомпонентные, состоящие из называемого апоферментом, и небелковой части, называемой простетической группой. Апофермент двухкомпонентных Ф называют также носителем, а простетическую группу – активной группой. Благодаря работам О. Варбурга, А. Теорелля, Ф Линена, Ф Липмана и Л. Лелуара установлено, что простетические группы многих Ф представляют собой производные витаминов или нуклеотидов. Т. о. была открыта важнейшая функциональная связь между Ф, витаминами и нуклеотидами, являющимися строительными "кирпичиками" нуклеиновых кислот.
Примером двухкомпонентного Ф является пируватдекарбоксилаза, катализирующая расщепление пировиноградной кислоты на двуокись и уксусный альдегид: 3 ® 3 + 2. Простетическая группа пируватдекарбоксилазы (тиаминнирофосфат) образована молекулой тиамина (витамина 1) и двумя остатками кислоты. Простетические группы ряда важных окислительно-восстановительных Ф – содержат производное амида никотиновой кислоты (ниацина), или же рибофлавина (витамина 2); в состав простетических группы т. н. пиридоксалевых ферментов, катализирующих перенос аминогрупп (–2) и декарбоксилирование и ряд др. превращений аминокислот, входит пиридоксальфосфат – производное витамина 6; активная группа Ф, катализирующих перенос остатков различных органических кислот (например, ацетила 3–), включает витамин пантотеновую кислоту. К двухкомпонентным Ф относятся также важные окислительные Ф – каталаза (катализирует реакцию разложения перекиси на воду и и пероксидаза (окисляет перекисями различные соединения, например полифенолы с образованием соответствующего хинона и воды). Каталитическое действие этих Ф может быть воспроизведено с помощью ионов трехвалентного Эти ионы обладают, однако, очень малой каталитической активностью, которая может быть усилена, если входит в состав гема. Хотя гем обладает уже значительным каталазным действием, его каталитическая активность все же в несколько миллионов раз меньше активности каталазы, в которой гем в качестве простетической группы этого Ф связан со специфическ им Гем обладает также слабым пероксидазным действием, однако это действие проявляется в полной мере только после соединения гема со специфическим в целостный Ф пероксидазу. Т. о., соединение простетической группы с приводит к резкому возрастанию ее каталитической активности. Вместе с тем от природы зависит не только каталитическая активность, но и специфичность действия Ф Прочность связи простетической группы и апофермента различна у разных Ф У некоторых Ф, например у катализирующих окисление различных субстратов путем отщепления эта связь является непрочной. Такие Ф легко диссоциируют (например, при диализе) и распадаются на простетическую группу и апофермент. Простетические группы, легко отделяющиеся от части Ф, называются коферментами.
Многие Ф содержат металлы, без которых Ф не активен. Эти металлы называются кофакторам и. Так, пероксидаза и каталаза содержат аскорбинатоксидаза, катализирующая окисление аскорбиновой кислоты, – алкогольдегидрогеназа, окисляющая спирты в соответствующие альдегиды, –
Специфичность и механизм действия ферментов. Действие Ф, в отличие от неорганических катализаторов, строго специфично и зависит от строения субстрата, на который Ф действует. Прекрасным примером такой зависимости служит катализируемая аргиназой реакция гидролитического расщепления аминокислоты аргинина на орнитин и мочевину:
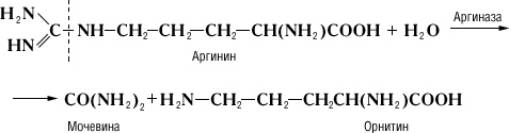 Однако аргиназа не расщепляет метилового эфира аргинина: Однако аргиназа не расщепляет метилового эфира аргинина:
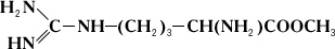
Дипептид, состоящий из остатков двух молекул аргинина, под действием аргиназы дает лишь половину теоретического количества мочевины. Очевидно, что, хотя расщепление аргинина происходит в месте, весьма отдаленном от карбоксильной () группы (показано пунктиром), необходимым условием действия аргиназы является ее соединение с карбоксильной группой аргинина. Поэтому замещение в карбоксильной группе на метильный остаток или же связывание карбоксильной группы со второй молекулой аргинина оказывают резкое влияние на действие аргиназы. Примеры специфичности действия Ф могут быть приведены при рассмотрении их стереохимической специфичности, т. е. действия Ф на стереоизомеры (см. Изомерия). Так, Ф, окисляющий природные L-аминокислоты, не действует на D-изомеры этих же аминокислот; Ф дипептидаза, гидролизирующий дипептиды, состоящие из остатков L-аминокислот, не действует на такие же дипептиды, состоящие из остатков D-аминокислот. Специфичность действия Ф послужила нем. ученому Э. Фишеру основанием для сравнения субстрата и Ф, который катализирует его превращение, с замком и соответствующим ему ключом. Стереохимическая специфичность Ф теснейшим образом связана с одной из основных особенностей живых организмов – их способностью к синтезу оптически активных органических соединений.
В образовании соединения между ферментом и субстратом – т. н. фермент-субстратного комплекса – принимают участие лишь некоторые функциональные группы молекулы Ф, образующие его активный центр. Так, например, в молекуле гидролизирующего химотрипсина, состоящего из 246 аминокислотных остатков, активный центр образован одним из остатков серина (химотрипсин относится к сериновым и двумя остатками гистидина, расположенными в удаленных друг от друга участках полипептидной цепи. Сближение этих функциональных групп активного центра происходит благодаря свойственной молекуле химотрипсина специфической пространственной (третичной) структуре. Ее нарушение в результате денатурации или каких-либо модификаций приводит к изменению или полной потере каталитической активности. В случае двухкомпонентных Ф в образовании фермент-субстратного комплекса принимают участие не только функциональные группы апофермента, но и простетическая группа. Так, при расщеплении пировиноградной кислоты пируватдекарбоксилазой субстрат связывается с частью молекулы тиамин-пирофосфата следующим образом:
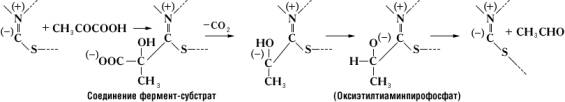
Исключительно высокая специфичность действия Ф объясняется их природой. Так, пиридоксалевые Ф, содержащие один и тот же кофермент (пиридоксальфосфат), могут принадлежать к различным классам и катализировать самые разнообразные реакции. Специфичность их действия зависит от природы апофермента.
Условия действия ферментов. Действие Ф зависит от ряда факторов, прежде всего от температуры и реакции среды (pH). Оптимальная температура, при которой активность Ф наиболее высока, находится обычно в пределах 40–50 °С. При более низких температурах скорость ферментативной реакции, как правило, снижается, а при температурах, близких к 0 °С, практически реакция полностью прекращается. При повышении температуры выше оптимальной скорость ферментативной реакции также снижается и, наконец, полностью прекращается. Снижение интенсивности действия Ф при повышении температуры сверх оптимальной объясняется главным образом начинающимся разрушением (денатурацией) входящего в состав Ф Поскольку в сухом состоянии денатурируются значительно медленнее, чем оводненные (в виде геля или раствора), инактивирование Ф в сухом состоянии происходит гораздо медленнее, чем в присутствии влаги. Поэтому сухие споры бактерий или сухие семена могут выдержать нагревание до гораздо более высоких температур, чем те же споры или семена в увлажненном состоянии.
Важнейшим фактором, от которого зависит действие Ф, как установил впервые С. Серенсен, является активная реакция среды – pH. Отдельные Ф различаются по оптимальной для их действия величине pH. Так, например, пепсин, содержащийся в желудочном соке, наиболее активен в сильнокислой среде (pH 1–2); трипсин – протеолитический Ф, выделяемый поджелудочной имеет оптимум действия в слабощелочной среде (pH 8–9); оптимум действия папаина – протеолитического Ф растительного происхождения – находится в слабокислой среде (pH 5–6).
Действие Ф зависит также от присутствия специфических активаторов и неспецифических или специфических ингибиторов. Так, энтерокиназа, выделяемая поджелудочной превращает неактивный трипсиноген в активный трипсин. Подобные неактивные Ф, содержащиеся в клетках и в секретах различных желез, называются проферментами. Многие Ф активируются в присутствии соединений, содержащих сульфгидрильную группу (–). К ним принадлежат аминокислота цистеин и трипептид глутатион, содержащийся в каждой живой клетке. Особенно сильное активирующее действие глутатион оказывает на некоторые протеолитические и окислительные Ф Неспецифическое угнетение (ингибирование) Ф происходит под действием различных веществ, дающих с нерастворимые осадки или блокирующих в них какие-либо группы (например, -группы). Существуют более специфические ингибиторы Ф, угнетение которыми каталитических функций основано на специфическом связывании этих ингибиторов с определенными группировками в активном центре Ф Так, окись () специфически ингибирует ряд окислительных Ф, содержащих в активном центре или Вступая в соединение с этими металлами, она блокирует активный центр Ф и вследствие этого он теряет свою активность. Различают обратимое и необратимое ингибирование Ф В случае обратимого ингибирования (например, действие малоновой кислоты на сукцинатдегидрогеназу) активность Ф восстанавливается при удалении ингибитора диализом или иным способом. При необратимом ингибировании действие ингибитора, даже при очень низких его концентрациях, усиливается со временем и в конце концов наступает полное торможение активности Ф Ингибирование Ф может быть конкурентным и неконкурентным. При конкурентном ингибировании ингибитор и субстрат конкурируют между собой, стремясь вытеснить один другого из фермент-субстратного комплекса. Действие конкурентного ингибитора снимается высокими концентрациями субстрата, в то время как действие неконкурентного ингибитора в этих условиях сохраняется. Действие на Ф специфических активаторов и ингибиторов имеет большое значение для регулирования ферментативных процессов в организме.
Классификация и номенклатура ферментов. По рекомендации Международного биохимического союза, Ф разделяют на 6 классов: 1) оксидоредуктазы, 2) трансферазы, 3) гидролазы, 4) лиазы, 5) изомеразы, 6) лигазы. Рекомендована следующая нумерация Ф Шифр (индекс) каждого Ф содержит 4 числа, разделенных точками. Первая цифра указывает класс, вторая – подкласс, третья – подподкласс, четвертая – порядковый номер в данном подподклассе. Так, Ф аргиназа, расщепляющий аргинин на орнитин и мочевину, имеет шифр 3.5.3.1, т. е. относится к классу гидролаз, подклассу Ф, действующих на непептидные С–-cвязи, и подподклассу Ф, расщепляющих эти связи в линейных (не циклических) соединениях.
Класс оксидоредуктаз включает Ф, катализирующие окислительно-восстановительные реакции, и разделяется на 14 подклассов в зависимости от природы той группы в молекуле субстрата, которая подвергается окислению (спиртовая, альдегидная, кетонная и т.д.). Подподклассы оксидоредуктаз индексируются в зависимости от типа участвующего в реакции акцептора (электронов) – кофермента, цитохрома, молекулярного и т.д. Т. о., первые три цифры шифра определяют тип Ф, так, например, 1.2.3 обозначают оксидоредуктазу, действующую на альдегид с молекулярным в качестве акцептора электронов. Класс трансфераз, объединяющий Ф, катализирующие реакции переноса групп, подразделяется на 8 подклассов в зависимости от природы переносимых групп, которыми могут быть одноуглеродные или гликозильные остатки, или содержащие серу группы и т.д. У трансфераз третья цифра характеризует тип переносимых групп (например, одноуглеродная группа может быть метилом, карбоксилом, формилом и т.д.). К гидролазам принадлежат Ф, катализирующие гидролитическое расщепление различных соединений; разделяются на 9 подклассов в зависимости от типа гидролизуемой связи – сложноэфирной, гликозидной и т.д. Третья цифра у гидролаз уточняет тип гидролизуемой связи. Лиазы – Ф, отщепляющие от субстрата ту или иную группу (негидролитическими путями) с образованием двойной связи или, наоборот, присоединяющие группы к двойным связям. У лиаз 5 подклассов, вторая цифра шифра обозначает тип подвергающейся разрыву связи ( – – и т.д.), а третья – тип отщепляемой группы. Изомеразы, катализирующие реакции изомеризации, разделяются на 5 подклассов в зависимости от типа катализируемой реакции; третья цифра шифра детализирует характер превращения субстрата. Лигазами (или синтетазами) называются Ф, которые катализируют соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле кислоты (АТФ) или аналогичного трифосфата. Первая цифра шифра лигаз обозначает тип вновь образуемой связи ( – – и т.д.), а вторая – природу образующегося соединения.
Классификация и номенклатура Ф, кроме шифра, включает также систематические и тривиальные (рабочие) названия. Так, например, систематическое название карбоксилаза 2-оксокислот соответствует уже упоминавшемуся тривиальному название пируватдекарбоксилаза, а систематическое название L-apгинин – амидиногидролаза – рабочему название аргиназа.
Регуляция ферментативных процессов. Действие Ф в организме осуществляется путем регуляции их синтеза и активности. Свойственный данному организму набор Ф определяется его генетической природой. Однако он может изменяться под влиянием различных внутренних и внешних факторов – мутаций, действия ионизирующей радиации, состава газовой среды, условий питания и т.д. Так, в результате мутаций возникают т. н. "молекулярные болезни" (например, алкаптонурия). При этом наследственном заболевании у больных с мочой выделяется гомогентизиновая кислота, образующаяся в результате превращений аминокислоты тирозина. Гомогентизиновая кислота накапливается в организме и выделяется с мочой вследствие того, что у больных алкаптонурией утеряна способность к синтезу двух Ф, катализирующих ее дальнейшее окисление, – параоксифенилпируватоксидазы и оксидазы гомогентизиновой кислоты. Влияние условий питания организма на его ферментный аппарат особенно наглядно прослеживается у микроорганизмов. Например, кишечная палочка при росте на питательной среде, содержащей синтезирует только следы b-галактозидазы. В присутствии же различных b-галактозидов образуются значительные количества этого Ф – до 6–7% от всех содержащихся в клетке Ф, новообразование или усиление синтеза которых происходит под влиянием какого-либо соединения, называются индуцируемыми ферментами. Под влиянием др. соединений может происходить подавление синтеза Ф, называемое репрессией. В животном организме индукция и репрессия синтеза Ф осуществляется не только под влиянием соответствующих субстратов и метаболитов, но и под влиянием гормонов. Так, синтез принимающей участие в синтезе в печени, индуцируется гормонами тироксином и кортизоном, но репрессируется инсулином. Общая теория индукции и репрессии биосинтеза на генетическом уровне дана французскими учеными Ф Жакобом и Ж. Моно (см. Оперон). В одном организме один и тот же Ф может быть представлен различными молекулярными формами. Такие разнообразные формы Ф, катализирующие одну и ту же реакцию, но различающиеся по физическим, и иммунологическим свойствам, называются изоферментами. Синтез изоферментов определяется генетическими факторами, но может изменяться под влиянием условий существования организма. Т. о., факторы, от которых зависят концентрация и активность Ф в организме, так же разнообразны, как и условия его существования. Это прежде всего водный, газовый, температурный, кислотный и световой режим среды, а также концентрация субстратов и различных кофакторов, необходимых для действия Ф, наличие активаторов и ингибиторов, концентрации метаболитов и, наконец, у высших многоклеточных организмов это нервная и гормональная регуляция ферментативной активности.
Примером влияния условий существования организма на активность Ф может служить Пастера эффект – прекращение брожения под действием Активность многих Ф регулируется по аллостерическому принципу. У таких Ф имеется т. н. аллостерический центр, присоединяясь к которому определенный метаболит – эффектор вызывает изменение структуры активного центра, вследствие чего активность Ф снижается или повышается.
Некоторые Ф находятся в клетке в виде многоферментных комплексов. В таких многоферментных ансамблях активность каждого отдельного Ф строго координирована и регулируется др. Ф, входящими в состав данного комплекса. Примером многоферментного комплекса может служить пируватдегидрогеназа, состоящая из 16 молекул пируватдекарбоксилазы, 8 молекул дигидролипоилдегидрогеназы и 4 агрегатов липоатацетилтрансферазы, каждая из которых состоит из 16 субъединиц. Решающую роль в регуляции активности Ф в клетке играют различные субклеточные структуры – митохондрии, микросомы, лизосомы и т.д., и мембраны, отделяющие их от цитоплазмы. Многие Ф вмонтированы в этих мембранах в виде многоферментных ансамблей.
Практическое значение ферментов. Ферментативные процессы являются основой многих производств: хлебопечения, виноделия, пивоварения, сыроделия, производства спирта, чая, уксуса. С начала 20 в. по предложению япон. ученого Д. Такамине в спиртовой и др. отраслях промышленности началось применение ферментных препаратов, получаемых из плесневых грибов или бактерий. В ряде стран этот способ широко используется для осахаривания с помощью амилаз крахмалистого сырья с целью получения или его сбраживания на спирт. Концентрированные амилолитические препараты Ф из плесневых грибов при добавке в тесто приводят к улучшению качества хлеба и ускорению технологического процесса. Препараты протеолитических Ф, получаемых из микроорганизмов, употребляются в кожевенной промышленности для удаления волос и мягчения сырья, а в сыродельной промышленности – для замены дефицитного сычужного фермента (реннина). Препараты микробных пектолитических Ф широко используют при производстве соков (выход плодового сока повышается на 10–20%). Все большее применение очищенные ферментные препараты находят в медицине. В научных исследованиях и в клинической практике высокоочищенные ферментные препараты служат в качестве специфических средств биохимического анализа (см. Ферментативные методы анализа). Весьма перспективно применение т. н. иммобилизованных Ф, которые связываются каким-либо носителем, образующим с данным Ф нерастворимый комплекс. При подборе соответствующего носителя можно получить иммобилизованный Ф с высокой активностью, устойчивый по отношению к денатурирующим агентам. Колонка, заполненная иммобилизованным Ф, может быть многократно использована для проведения соответствующей реакции. Иммобилизованные Ф находят все более широкое применение в аналитической практике и биохимической технологии.
Лит.: Ф, М., 1964; Диксон М., Уэбб Э., Ф, пер. с англ., М., 1966; Номенклатура ферментов, пер. с англ., М., 1966; Бернхард С., Структура и функция ферментов, пер. с англ., М., 1971; Структура и функция ферментов, в. 1–2, М., 1972–73; Фениксова Р. В., Биохимические основы получения и применения ферментных препаратов, в кн.: Техническая биохимия, М., 1973; Кретович В. Л., Введение в энзимологию, 2 изд., М,, 1974; Аллостерические ферменты, М., 1975; Ф медицинского назначения, Л., 1975; Ферментные препараты в пищевой промышленности, М., 1975; Advances in enzymology and related areas of molecular biology, v. 1–43, . ., 1941–75; Methods in enzymology, v. 1–36, . ., 1955–75.
В. Л. Кретович.
|
Для поиска, наберите искомое слово (или его часть) в поле поиска
|
|
 |
 |
 |
|
|
|
Новости 21.11.2024 12:23:12
|
|
|
|
|
|
|
|
|
|
|


