
|

|
Большая Советская Энциклопедия (цитаты)
|

|
|
|
|
Дыхание | Дыхание (далее Д) совокупность процессов, которые обеспечивают поступление в организм и выделение из него углекислого газа (внешнее Д) и использование клетками и тканями для окисления органических веществ с освобождением содержащейся в них энергии, необходимой для жизнедеятельности (тканевое дыхание, клеточное Д). Бескислородный путь освобождения энергии свойствен только небольшой группе организмов - так называемым анаэробам (см. Брожение); в ходе эволюции освобождение энергии в результате Д стало у подавляющего большинства организмов главным процессом, а анаэробные реакции сохранились в основном как промежуточные этапы обмена веществ.
Д животных и человека. У простейших, губок, кишечнополостных и некоторых др. организмов О2 диффундирует непосредственно через поверхность тела. С усложнением организации и увеличением размеров тела появляются специальные дыхания органы, а также система кровообращения, в которой циркулирует жидкость - кровь или гемолимфа, содержащая вещества, способные связывать и переносить 2 и 2 (см. Гемоглобин). У насекомых 2 поступает в ткани из системы воздухоносных трубочек - трахей. У водных животных, использующих растворенный в воде 2, органами Д служат жабры, снабженные богатой сетью кровеносных сосудов. В этом случае 2, растворенный в воде, диффундирует в кровь, циркулирующую в сосудах жаберных щелей. У многих рыб значительную роль играет кишечное Д, при котором воздух заглатывается и 2 поступает в кровь через кровеносные сосуды кишечника; некоторую роль в Д рыб играет также плавательный пузырь; у многих обитающих в воде животных обмен газов (главным образом СО2) происходит и через кожу. У сухопутных животных внешнее Д обеспечивается преимущественно легкими. У земноводных и многих др. животных наряду с этим функционирует кожное Д У птиц существенное значение имеют сообщающиеся с легкими воздушные мешки, которые изменяются в объеме при летательных движениях и облегчают Д в полете. У земноводных и пресмыкающихся воздух в легкие нагнетается движениями мышц дна рта. У птиц, млекопитающих и человека внешнее Д обеспечивается ритмической работой дыхательных мышц (главным образом диафрагмы и межреберных мышц), координируемой нервной системой. При сокращении этих мышц объем грудной клетки увеличивается и происходит растяжение находящихся в ней легких; поэтому возникает разность между атмосферным и внутрилегочным давлением и воздух поступает в легкие (вдох). Выдох может быть пассивным - за счет спадения растянутой во время вдоха грудной клетки, а вслед за ней и легких; активный выдох обусловлен сокращением некоторых групп мышц. Количество воздуха, поступающее в легкие за 1 вдох, называется дыхательным объемом (см. Легочные объемы).
При Д дыхательная мускулатура преодолевает эластичное сопротивление, связанное с упругостью грудной клетки, тягой легких и поверхностным натяжением альвеол. Последнее, однако, значительно снижается под влиянием поверхностно активного вещества, вырабатываемого клетками альвеолярного эпителия; поэтому альвеолы при выдохе не спадаются, а при вдохе легко расширяются. Чем выше эластичное сопротивление, тем труднее растягиваются грудная клетка и легкие; при глубоком Д работа дыхательной мускулатуры, затрачиваемая на его преодоление, резко возрастает. Неэластичное сопротивление Д обусловлено главным образом трением воздуха при его движении по носовым ходам, гортани, трахее и бронхам. Оно зависит от скорости потока воздуха во время Д и от его характера. При спокойном Д поток близок к ламинарному (линейному) в прямых участках воздухоносных путей и к турбулентному (вихревому) в местах разветвления или сужения. С увеличением скорости потока (при форсированном Д) турбулентность возрастает и для продвижения воздуха требуется более высокая разность давлений, а следовательно, и увеличение работы дыхательных мышц. Неравномерное распределение сопротивления движению воздуха по дыхательным путям приводит к тому, что поступление воздуха в разные группы легочных альвеол происходит неравномерно; такая разница в вентиляции особенно значительна при легочных заболеваниях.
Количество воздуха, вентилирующее легкие в 1 мин, называется минутным объемом дыхания (МОД). МОД равен произведению дыхательного объема на частоту Д (число дыхательных движений в 1 мин, равное у человека примерно 15-18) и составляет у взрослого человека в покое 5-8 л/мин. Только часть МОД (около 70%) участвует в обмене газов между вдыхаемым и альвеолярным воздухом, эту часть называют объемом альвеолярной вентиляции; остальная часть МОД используется для "промывания" так называемого мертвого, или вредного, пространства дыхательных путей, в котором к началу выдоха сохраняется наружный воздух, заполнивший его в конце предшествовавшего вдоха (объем мертвого пространства около 160 мл). Вентиляция альвеол обеспечивает постоянство состава альвеолярного воздуха. Парциальное давление 2(p2) и 2 (p2) в альвеолярном воздухе колеблется в очень узких пределах и составляет для О2 около 13 кн/м2 (100 мм рт. cт.) и для СО2 около 5,4 кн/м2 (40 мм рт. ст.).
Обмен газов между альвеолярным воздухом и венозной кровью, поступающей в капилляры легких, осуществляется через альвеоло-капиллярную мембрану, общая поверхность которой очень велика (у человека около 90 м2). Диффузия 2 в кровь обеспечивается разностью парциальных давлений 2 в альвеолярном воздухе и в венозной крови (8-9 кн/м2, или 60-70 мм рт. ст.). 2, приносимый кровью из тканей в связанной форме (бикарбонаты, соли кислоты и карбгемоглобин), освобождается в капиллярах легких при участии фермента карбоангидразы и диффундирует из крови в альвеолы; разность p2 между венозной кровью и альвеолярным воздухом составляет около 7 мм рт. ст. Способность альвеолярной стенки пропускать 2 и 2, так называемая диффузионная способность легких, очень велика: в покое она составляет в 1 мин примерно 30 мл 2 на 1 мм разности p2 между альвеолярным воздухом и кровью; для 2 эта величина во много раз больше. Поэтому парциальное давление газов в оттекающей из легких артериальной крови успевает приблизиться к их давлению в альвеолярном воздухе. Переход 2 в ткани и удаление из них 2 также происходят путем диффузии, т.к. p2 в тканевой жидкости 2,7-5,4 кн/м2 (20-40 мм рт. ст.), а в клетках еще ниже, а p2 в клетках может достигать 60 мм рт. cт. (см. рис.).
Потребление 2 клетками и тканями и образование ими 2, что составляет сущность тканевого, или клеточного, Д, - одна из основных форм диссимиляции, осуществляющейся у животных и растений в принципе одинаково. Высокое потребление 2 характерно для тканей почек, коры больших полушарий головного мозга, сердца. В результате окислительно-восстановительных реакций тканевого Д освобождается энергия, расходуемая на все жизненные проявления. Процесс этот осуществляется в митохондриях и складывается из дегидрирования субстратов Д - углеводов и продуктов их расщепления, жиров и жирных кислот, аминокислот и продуктов их дезаминирования. Субстраты Д поглощают 2 и служат источником 2 (отношение -
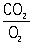
называется дыхательным коэффициентом). Энергия, освобождающаяся при окислении органических веществ, не используется тканями непосредственно, т.к. около 70% ее расходуется на образование АТФ - одной из аденозинфосфорных кислот, последующее ферментативное расщепление которой обеспечивает энергетические потребности тканей, органов и организма в целом (см. Окисление биологическое, Окислительное фосфорилирование). Т. о., с биохимической точки зрения Д - это превращение энергии углеводов и др. веществ в энергию макроэргических фосфатных связей.
Постоянство p2 и p2 в альвеолярном воздухе, а стало быть, и в артериальной крови может поддерживаться лишь при условии, если альвеолярная вентиляция соответствует скорости потребления организмом 2 и образования 2, т. е. уровню обмена веществ. Это условие обеспечивается благодаря совершенным механизмам регуляции Д Управление частотой и глубиной Д осуществляется рефлекторным путем. Так, повышение p2 и снижение p2 в альвеолярном воздухе и в артериальной крови возбуждают хеморецепторы синокаротидной и кардиоаортальной зон, что приводит к возбуждению дыхательного центра и увеличению МОД Согласно классическим представлениям, повышение p2 в артериальной крови, омывающей дыхательный центр, также возбуждает его и вызывает увеличение МОД Т. о., регуляция Д по отклонению p2 и p2 в артериальной крови, осуществляемая по типу обратной связи, обеспечивает оптимальный МОД Однако в ряде случаев, например при мышечной работе, МОД увеличивается до наступления в обмене веществ сдвигов, которые приводят к изменениям в газовом составе крови. Это усиление вентиляции обусловлено сигналами, поступающими в дыхательный центр от рецепторов двигательного аппарата, двигательной зоны коры больших полушарий мозга, а также условными рефлексами на различные сигналы, связанные с привычной работой и ее обстановкой. Т. о., управление Д осуществляется сложной самообучающейся системой не только по принципу регулирования по отклонению, но и по сигналам, предупреждающим о возможных отклонениях. Смена вдоха и выдоха обеспечивается системой взаимодополняющих механизмов. Во время вдоха в дыхательный центр по волокнам блуждающих нервов поступают импульсы от рецепторов растяжения, находящихся в легких. При достижении легкими определенного объема эта импульсация тормозит клетки дыхательного центра, возбуждение которых вызывает вдох. При выключении нервных путей, обеспечивающих поступление импульсов в дыхательный центр, ритмичность Д сохраняется благодаря автоматизму центра, однако характер ритма резко отличается от нормального. При нарушениях Д и механизмов его регуляции возникают изменения газового состава крови (см. Гипоксия).
Методы исследования Д разнообразны. В физиологии труда и спорта, клинической медицине широко применяют регистрацию глубины и частоты дыхательных движений, измерения газового состава выдыхаемого воздуха, артериальной крови, плеврального и альвеолярного давления. См. также Газообмен.
Лит.: Сеченов И. М., Избр. труды, М., 1935; Холден Дж. и Пристли Дж., Д, пер. с англ., М.-Л., 1937; Маршак М. Е., Регуляция дыхания у человека, М., 1961; Физиология человека, М., 1966; Comroe J. Н., Physiology of respiration, Chi., 1966; Dejours ., Respiration, Oxf., 1966.
Л. Л. Шик.
Д растений. Д присуще всем органам, тканям и клеткам растения. Об интенсивности Д можно судить, измеряя количество выделяемого тканью 2 либо поглощаемого ею 2. Более интенсивно дышат молодые, быстро растущие органы и ткани растений. Наиболее активно Д репродуктивных органов, затем листьев; слабее Д стеблей и корней. Теневыносливые растения дышат слабее светолюбивых. Для высокогорных растений, адаптированных к пониженному парциальному давлению 2, характерна повышенная интенсивность Д Очень активно Д плесневых грибов, бактерий. Д усиливается с повышением температуры (на каждые 10°С - примерно в 2-3 раза), прекращаясь при 45-50°С. В тканях зимующих органов растений (почки лиственных деревьев, иглы хвойных) Д продолжается (с резко сниженной интенсивностью) и при значительных морозах. Д стимулируют механические и раздражения (поранения, некоторые яды, наркотики и т.п.). Закономерно изменяется Д в ходе развития растения и его органов. Сухие (покоящиеся) семена дышат очень слабо; при набухании и последующем прорастании семян Д усиливается в сотни и тысячи раз. С окончанием периода активного роста растений Д их тканей ослабевает, что связано с процессом старения протоплазмы. При созревании семян, плодов интенсивность Д уменьшается.
Согласно теории советского биохимика А. Н. Баха, процесс Д, т. е. окисление углеводов, жиров, осуществляется при помощи окислительной системы клетки в два этапа: 1) активирование 2 воздуха путем его присоединения к содержащимся в живой клетке ненасыщенным, способным самопроизвольно окисляться соединениям (оксигеназам) с образованием перекисей; 2) активирование последних с освобождением способного окислять трудно окисляемые органические вещества. По теории дегидрирования русского ботаника В. И. важнейшее звено Д - активация субстрата, осуществляемая дегидрогеназами. Обязательный участник сложной цепи процессов Д - вода, которой вместе с субстрата используется для восстановления самоокисляющихся соединений - так называемых дыхательных пигментов. 2, выделяющийся при Д, образуется без участия воздуха, т. е. анаэробно. воздуха идет на окисление дыхательных превращающихся при этом в дыхательные пигменты. Дальнейшее развитие теория Д получила в исследованиях советского ботаника С. П. Костычева, согласно которым первые этапы аэробного Д аналогичны процессам, свойственным анаэробам. Превращения образующегося при этом промежуточного продукта могут идти с участием что свойственно аэробам. У анаэробов же эти превращения идут без участия молекулярного По современным представлениям, процесс окисления, который составляет основу Д, заключается в потере веществом электрона. Способность присоединять или отдавать электроны зависит от величины окислительного потенциала соединения. обладает самым высоким окислительным потенциалом и, следовательно, максимальной способностью присоединять электроны. Однако потенциал сильно отличается от потенциала дыхательного субстрата. Поэтому роль промежуточных переносчиков электронов от дыхательного субстрата к выполняют специфические соединения. Попеременно окисляясь и восстанавливаясь, они образуют систему переноса электронов. Присоединив к себе электрон от менее окисленного компонента, такой переносчик восстанавливается и, отдавая его следующему компоненту с более высоким потенциалом, окисляется. Так электрон передается от одного звена дыхательной цепи к другому и, в конце концов, Таков заключительный этап Д
Все эти процессы (активация перенос электрона по цепи на осуществляются главным образом в митохондриях благодаря разветвленной системе окислительно-восстановительных ферментов (см. Цитохромы). По пути следования к электроны, мобилизуемые первоначально от молекулы органического вещества, постепенно отдают заключенную в них энергию, которую клетка запасает в форме соединений, главным образом АТФ.
Благодаря совершенным механизмам запасания и использования энергии процессы энергообмена в клетке идут с очень высоким кпд, пока недостижимым в технике. Биологическая роль Д не исчерпывается использованием энергии, заключенной в окисляемой органической молекуле. В ходе окислительных превращений органических веществ образуются активные промежуточные соединения - метаболиты, которые живая клетка использует для синтеза специфических составных частей своей протоплазмы, образования ферментов и др. Всем этим определяется центральное место, занимаемое Д в комплексе процессов обмена веществ живой клетки. В Д скрещиваются и увязываются процессы обмена нуклеиновых кислот, углеводов, жиров и др. компонентов протоплазмы.
Лит.: Костычев С. П., Физиология растений, 3 изд., т. 1, М.-Л., 1937; Бах А. Н., Собр. трудов по химии и биохимии, М., 1950; Таусон В. О., Основные положения растительной биоэнергетики, М.-Л., 1950; Джеймс В. О., Д растений, пер. с англ., М., 1956; В. И., Избр. труды, М., 1960; Михлин Д М., Биохимия клеточного дыхания, М., 1960; Сент-Дьердьи А., Биоэнергетика, пер. с англ., М., 1960; Рубин Б. А., Ладыгина М. Е., Энзимология и биология дыхания растений, М., 1966; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Рубин Б. А., Курс физиологии растений, 3 изд., М., 1971; Кретович В. Л., Основы биохимии растений, М., 1971.
Б. А. Рубин.
|
Для поиска, наберите искомое слово (или его часть) в поле поиска
|
|
 |
 |
 |
|
|
|
Новости 21.11.2024 12:16:15
|
|
|
|
|
|
|
|
|
|
|


