
|

|
Большая Советская Энциклопедия (цитаты)
|

|
|
|
|
Фотосинтез | Фотосинтез (далее Ф) (от фото... и синтез), образование высшими растениями, водорослями, фотосинтезирующими бактериями сложных органических веществ, необходимых для жизнедеятельности как самих растений, так и всех др. организмов, из простых соединений (например, углекислого газа и воды) за счет энергии света, поглощаемой и др. фотосинтетическими пигментами. Один из важнейших биологических процессов, постоянно и в огромных масштабах совершающийся на нашей планете. В результате Ф растительность земного шара ежегодно образует более 100 млрд. т органического веществ (около половины этого количества приходится на долю Ф растений морей и океанов), усваивая при этом около 200 млрд. т 2 и выделяя во внешнюю среду около 145 млрд. т свободного Полагают, что благодаря Ф образуется весь атмосферы. Ф – единственный биологический процесс, который идет с увеличением свободной энергии системы; все остальные (за исключением хемосинтеза) осуществляются за счет потенциальной энергии, запасаемой в продуктах Ф Количество энергии, ежегодно связываемой фотосинтезирующими организмами океана и суши (около 3×1021 дж), во много раз больше той энергии, которая используется человечеством (около 3×1020 дж).
Историческая справка. Начало исследованию Ф положено работами Дж. Пристли, Ж. Сенебье, Н. Соссюра, Я. Ингенхауза, Ю. Майера, в которых постепенно выяснилось, что растения на свету усваивают из воздуха углекислый газ, выделяют образуют в результате этого органические вещества, запасая в них энергию солнечного света. Во 2-й половине 19 в. К. А. Тимирязев показал, что энергия солнечного света вводится в цепь фотосинтетических превращений через зеленый пигмент растений – хлорофилл: спектр действия Ф соответствует спектру поглощения света и интенсивность Ф увеличивается с увеличением интенсивности света. В 1905 английским ученый Ф Блекман обнаружил, что Ф состоит из быстрой световой реакции и более медленной – темновой. Биохимическое доказательство существования световой и темновой фаз были получены лишь в 1937 английским исследователем Р. Хиллом. Крупный вклад в изучение темновой и световой стадий Ф внесли также нем. биохимик и физиолог О. Варбург, амер. биохимик Х. Гафрон. В 1931 амер. микробиолог К. Нил показал, что фототрофные бактерии осуществляют Ф без выделения О2, т.к. при ассимиляции СО2 окисляют сероводород, тиосульфат и др. субстраты. Так было положено начало представлению о Ф как окислительно-восстановительном процессе, где восстановление 2 осуществляется при одновременном окислении донора В 1941 сов. учеными А. П. Виноградовым и М. В. Тейц, а также американскими исследователями Э. Рубеном и др. установлено, что источником выделяющегося в процессе Ф высших растений и водорослей, является вода, а не 2, как считали ранее.
Начиная с 1-й четверти 20 в. важные работы выполнены по изучению физиологии и экологии Ф (В. В. Сапожников, С. П. Костычев, В. Н. Любименко, А. А. Ничипорович, О. В. Заленский и многие др.). С середины 20 в. изучению Ф способствовало создание новых методов исследования (газовый анализ, изотопные методы, спектроскопия, электронная микроскопия и др.). Эти методы позволили разработать представления о тонких механизмах участия в Ф (А. Н. Теренин, А. А. Красновский, американские ученые Е. Рабинович, В. Кок, У. Арнолд, Р. Клейтон, Дж. Франк, франц. исследователь Дж. Лаворель); об окислительно-восстановительных реакциях Ф и о существовании двух фотохимических реакций Ф (английский фитофизиолог Р. Хилл, С. Очоа, амер. исследователи В. Вишняк, Р. Эмерсон, Френч, голландский ученый Л. Дейсенс); о фотосинтетическом (Д. Арнон); о путях превращения (М. Калвин, амер. ученые Дж. Бассам, Э. Бенсон, австралийские исследователи М. Хетч и К. Слэк); о механизме разложения воды (В. Кок, французские ученые А. и П. Жолио, советский ученый В. М. Кутюрин и др.).
Характерные черты фотосинтеза высших зеленых растений, водорослей и фотосинтезирующих бактерий. В реакциях Ф у высших зеленых растений, водорослей (многоклеточных – зеленых, бурых, красных, а также одноклеточных – эвгленовых, динофлагеллят, диатомовых) донором и источником выделяемого служит вода, а основным акцептором и источником – углекислый газ. При использовании в Ф только 2 и 2 образуются углеводы. Но в процессе Ф растения образуют не только углеводы, но и содержащие и серу аминокислоты, а также пигменты и др. соединения. Акцепторами (наряду с 2) и источниками и серы в этом случае служат нитраты (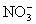 ) и сульфаты ( ) и сульфаты (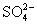 ). Фирующие бактерии не выделяют и не используют молекулярный (большинство из них облигатные, т. е. обязательные анаэробы). Вместо воды в качестве доноров электронов эти бактерии используют либо неорганические соединения (сероводород, тиосульфат, газообразный либо органические вещества (молочную кислоту, изопропиловый спирт). Источником в большинстве случаев является также 2, но наряду с этим и некоторые органические соединения (например, ацетат). Т. о., Ф у разных организмов может протекать с использованием различных доноров (ДН2), акцепторов (А) электронов и и может быть представлен схематически обобщенным уравнением: ). Фирующие бактерии не выделяют и не используют молекулярный (большинство из них облигатные, т. е. обязательные анаэробы). Вместо воды в качестве доноров электронов эти бактерии используют либо неорганические соединения (сероводород, тиосульфат, газообразный либо органические вещества (молочную кислоту, изопропиловый спирт). Источником в большинстве случаев является также 2, но наряду с этим и некоторые органические соединения (например, ацетат). Т. о., Ф у разных организмов может протекать с использованием различных доноров (ДН2), акцепторов (А) электронов и и может быть представлен схематически обобщенным уравнением:
Д×Н2 + А 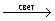 AH2 + Д, AH2 + Д,
где AH2 – продукты Ф
Структурные особенности фотосинтетического аппарата. Высокая эффективность Ф высших зеленых растений обеспечивается совершенным фотосинтетическим аппаратом, основа которого – внутриклеточные органеллы – хлоропласты (в клетке зеленого листа их 20–100). Они окружены двуслойной мембраной. Внутренний слой ее построен из уплощенных мешочков или пузырьков, называемых тилакоидами, которые часто упакованы в стопки, составляют граны, соединенные между собой одиночными межгранными тилакоидами. Тилакоиды состоят из собственно фотосинтетических мембран, представляющих собой биомолекулярные липидные слои и мозаично вкрапленные в них липопротеидопигментные комплексы, образующие фотохимически активные центры, и содержат также специальные компоненты, участвующие в транспорте электронов и образовании (АТФ). Часть находящаяся между тилакоидами строма, содержит ферменты, катализирующие темновые реакции Ф (например, превращение серы, биосинтез углеводов и В строме откладывается образуемый при Ф крахмал. имеют собственные ДНК, РНК, рибосомы, синтезирующие и обладают некоторой генетической автономностью, но находятся под общим контролем ядра. фотосинтезирующие бактерии и большинство водорослей не имеют Фотосинтетический аппарат большинства водорослей представлен специализированными внутриклеточными органеллами – хроматофорами, а фотосинтезирующих бактерий и сине-зеленых водорослей – тилакоидами (мембраны их содержат пигмент бактерио- или бактериовиридин, а также др. компоненты реакций Ф), погруженными в периферические слои цитоплазмы.
Фаза первичных превращений и запасания энергии в процессе Ф В основе Ф растений лежит окислительно-восстановительный процесс, в котором 4 электрона (и протона) поднимаются от уровня окислительно-восстановительного потенциала, соответствующего окислению воды (+ 0,8 в) до уровня, соответствующего восстановлению 2 с образованием углеводов (– 0,4 в). При этом увеличение свободной энергии реакции восстановления 2 до уровня углеводов составляет 120 ккал/моль, а суммарное уравнение Ф выражается как:
2 + 2  (2) + 2 + 120 ккал/моль. (2) + 2 + 120 ккал/моль.
Энергия моля квантов (Эйнштейна) красной части спектра составляет около 40 ккал/моль. Т. о., для Ф, идущего в соответствии с приведенным уравнением, было бы достаточно поглощения энергии 3 квантов на молекулу 2 (или на выделение молекулы 2). Однако в окислительно-восстановительной реакции от воды к 2 должны быть перенесены 4 электрона, причем перенос каждого из них осуществляется в ходе двух последовательных фотохимических реакций. Поэтому квантовый расход при оптимальных условиях составляет 8–12 квантов на молекулу 2, а максимальная эффективность преобразования энергии красного света – около 30° %. В полевых условиях вследствие неполного поглощения света, энергетических затрат на дыхание и др. потерь, а также ограниченности вегетационного периода эффективность усвоения солнечной энергии с.-х. растениями в умеренных широтах обычно не выше 0,5–1,3%. Сопоставление этих цифр c теоретическим максимальным значением указывает на существование значительных резервов, которые могут быть использованы в будущем. Для некоторых культур с.-х. растений удается в специальных условиях повысить энергетическую эффективность до 5–6% и даже выше (при культивировании водорослей до 7–10%).
Ни 2, ни вода непосредственно не поглощают свет, посредником во взаимодействии этих соединений с квантами служит а, включенный в структуру или и образующий функциональные фотосинтетические единицы, состоящие из нескольких сотен молекул пигмента и реакционных центров. Основная часть сопровождающих пигментов ( b, каротиноиды, фикобилины и др. и коротковолновые формы а) выполняет функцию светособирающей антенны. При поглощении квантов их молекулы переходят в возбужденное состояние, которое путем миграции энергии передается на молекулу а, находящуюся в реакционном центре. Эффективность передачи энергии обусловлена близким расположением молекул, а также наличием нескольких агрегированных форм а, участвующих в формировании реакционных центров и образующих нисходящую лестницу энергетических уровней. Возможен полупроводниковый перенос электрона по агрегированному пигменту. В реакционном центре происходит основной акт Ф – разделение зарядов с последующим образованием первичного окислителя и первичного восстановителя. Существуют два типа центров (рис. 1), один из которых включен в пигментную фотосистему (ФС ), а др. – в фотосистему (ФС ). В фотореакции, связанной с разложением воды, участвует ФС : пигментом ее центра служит а с максимумом поглощения 680 нм, гипотетическим первичным восстановителем – Q (вероятно, цитохром), а первичным окислителем – сложный комплекс Z. Возбуждение пигментной молекулы центра 680 сопровождается разделением зарядов и образованием окисленного Z+, который участвует в окислении воды и выделении 2. Полагают, что в систему разложения воды, пока мало изученную, входят неизвестные ферменты, ионы и бикарбонат. Первичный восстановитель Q (проявляется по индукции флуоресценции) ФС передает электрон переносчикам (цитохромы b, f, пластохинон, пластоцианин) фотосинтетической электронной транспортной цепи к реакционному центру ФС . Пигментом этого центра служит а с максимумом поглощения 700 нм, первичным восстановителем – неидентифицированное вещество Х. Восстановленный Х передает электрон ферредоксину – который восстанавливает никотинамидадениндинуклеотидфосфат (НАДФ). Его восстановленная форма – НАДФ-Н запасает основную часть энергии света. Др. часть энергии электронного потока запасается в виде АТФ (фотосинтетическое который образуется на нисходящем участке переноса электронов между ФС и ФС (нециклическое фотофосфорилирование) или при круговом замыкании потока в ФС (циклическое фотофосфорилирование). возможно, происходит по хемиосмотическому механизму за счет электрического потенциала и градиента концентрации Н+, возникающих при индуцировании светом электронного потока в мембранных структурах тилакоидов. Экспериментально обнаружено, что освещение индуцирует электрический потенциал на мембране Описанное последовательное соединение двух фотореакций и наиболее вероятно, хотя обсуждается возможность параллельного соединения реакций. Предполагают, что фотосинтезирующие бактерии осуществляют Ф с участием лишь одной пигментной фотосистемы, однако этот вопрос нельзя считать решенным. Фотофизические и фотохимические стадии заканчиваются за 10-12–10-8 сек разделением зарядов и последующим образованием первичного окислителя и восстановителя. Границей первичных биофизических и биохимических процессов обычно считают появление первых стабильных продуктов – НАДФ-Н и АТФ Эти вещества ("восстановительная сила") используются затем в темповых процессах восстановления 2.
Ассимиляция углекислоты. Ассимиляция 2 происходит в процессе темновых реакций. Восстановлению при Ф подвергается не свободная молекула 2, а предварительно включенная в состав определенного органического соединения. В большинстве случаев акцептором 2 служит двукратно пятиуглеродный рибулозодифосфат (РДФ). Присоединяя 2, РДФ распадается на 2 молекулы фосфоглицериновой кислоты (ФГК). 2, включенный в молекулу ФГК, и является конечным звеном цепи, к которому направляются электроны, мобилизуемые Присоединив электрон, ФГК превращается в восстановленное соединение – фосфоглицериновый альдегид (в этом процессе участвуют АТФ и НАДФ-Н), который может рассматриваться как первый стабильный углеводный продукт Ф, содержащий уже в восстановленной (органической) форме. Дальнейшие превращения происходят в пентозофосфатном цикле и завершаются, с одной стороны, образованием РДФ, т. е. происходит регенерация первичного акцептора 2 (что делает цикл при наличии света и 2 непрерывно действующим), а с др. стороны – образованием продуктов Ф – углеводов.
Все, что было сказано выше, относится к т. н. С3-растениям, которые усваивают в Ф через цикл Калвина (рис. 2), акцептируют 2 на РДФ при помощи РДФ-карбоксилазы, образуя первые трехуглеродные продукты Ф – фосфоглицериновую кислоту и фосфоглицериновый альдегид. Некоторые травянистые, главным образом тропического происхождения, растения (например, тростник, кукуруза, сорго) образуют в качестве первых продуктов Ф не трех-, а четырехуглеродные соединения – щавелевоуксусную, яблочную и аспарагиновую кислоты. Путь автотрофной ассимиляции 2 через фосфоенолпировиноградную кислоту, или фосфоенолпируват (ФЕП), с образованием С4-дикарбоновых кислот получил название С4-пути усвоения а организмы – С4-растений. В листьях таких растений имеется два типа фотосинтезирующих клеток и Ф идет в две стадии. В клетках мезофилла листа происходит первичное акцептирование 2 на ФЕП с участием ФЕП-карбоксилазы, которая вовлекает 2 в реакции карбоксилирования даже при очень низких концентрациях 2 в окружающем воздухе. В результате карбоксилирования образуются щавелевоуксусная, яблочная и аспарагиновая кислоты. Из них две последние переходят в обкладочные клетки проводящих пучков листа, подвергаются там декарбоксилированию и создают внутри клеток высокую концентрацию 2, усваиваемую уже через РДФ-карбоксилазу в цикле Калвина. Это выгодно, во-первых, потому, что облегчает введение 2 в цикл Калвина через карбоксилирование РДФ при помощи РДФ-карбоксилазы, которая менее активна и требует для оптимальной работы более высоких концентраций 2, чем ФЕП-карбоксилаза. Кроме того, высокая концентрация 2 в обкладочных клетках уменьшает световое дыхание (фотодыхание) и связанные с ним потери энергии. Т. о. происходит высокоинтенсивный "кооперативный" Ф, свободный от излишних потерь в световом дыхании, от ингибирования и хорошо приспособленный к осуществлению Ф в атмосфере, бедной 2 и богатой 2.
Существуют и др. пути превращения 2 при Ф, в результате которых в клетке в разных соотношениях образуются различные органические кислоты, и т.п. Соотношения между этими группами соединений в растении зависят от интенсивности и качества света, вида растения и условий его развития (корневого питания, условий освещения и др.). Регулируя условия развития растений, можно управлять составом продуктов Ф и тем самым – химизмом растения в целом.
Роль фотосинтеза в биосфере. Наряду с Ф на Земле совершаются примерно равноценные по масштабам, но противоположные по направлению процессы окисления органических веществ и восстановленного при горении топливных материалов (каменный нефть, газ, торф, дрова и т.п.), при расходовании органических веществ живыми организмами в процессе их жизнедеятельности (дыхание, брожение), в результате которых образуются полностью окисленные соединения – углекислый газ и вода, и освобождается энергия. Затем с помощью энергии солнечной радиации углекислый газ, вода снова вовлекаются в процессы Ф Т. о., энергия солнечного света, используемая при Ф, служит движущей силой колоссального по размерам круговорота на Земле таких элементов, как В этот круговорот включаются и многие др. элементы: , , Р, , и др. За время существования Земли благодаря Ф важнейшие элементы и вещества прошли уже много тысяч циклов полного круговорота.
В предшествующие эпохи условия для Ф на Земле были более благоприятны в связи с сильным перевесом восстановительных процессов над окислительными. Постепенно огромные количества восстановленного в органических остатках оказались захороненными в недрах Земли, образовав громадные залежи горючих ископаемых. В результате этого в атмосфере сильно снизилось относительное содержание углекислого газа (до 0,03 объемных %) и повысилось содержание что существенно ухудшило условия для Ф
Следствием появления на Земле мира фотосинтезирующих растений и непрерывного новообразования ими больших количеств богатых энергией органических веществ явилось развитие мира гетеротрофных организмов (бактерий, грибов, животных, человека) – потребителей этих веществ и энергии. В результате (в процессе дыхания, брожения, гниения, сжигания) органические соединения стали окисляться и подвергаться разложению в таких же количествах, в каких образуют их высшие растения, водоросли, бактерии. На Земле установился круговорот веществ, в котором сумма жизни на нашей планете определяется масштабами Ф В текущем геологическом периоде (антропогеновом) размеры фотосинтетической продуктивности на Земле, вероятно, стабилизировались. Однако в связи с бурно возрастающим использованием продуктов Ф основным ее потребителем – человеком – приходится думать о предстоящем истощении горючих ископаемых, пищевых, лесных ресурсов и т.п. Недостаточна фотосинтетическая мощность современной растительности для регенерации атмосферы: растительность Земли не способна полностью усваивать весь углекислый газ (относительное содержание его в атмосфере за последние 100 лет медленно, но неуклонно возрастает), дополнительно поступающий в окружающую среду в результате бурно возрастающих масштабов добычи и сжигания горючих ископаемых.
При этом потенциальная фотосинтетическая активность растений используется далеко не полно. Проблема сохранения, умножения и наилучшего использования фотосинтетической продуктивности растений – одна из важнейших в современном естествознании и практической деятельности человека.
Ф и урожай. Один из путей повышения общей продуктивности растений – усиление их фотосинтетической деятельности. Например, чтобы сформировать урожай пшеницы в 40 ц/га, что составляет 100 т общей сухой биомассы, растения должны усвоить около 20 т 2, фотохимически разложить около 7,3 т 2, выделить во внешнюю среду около 13 т 2. Обычно за время вегетации растений в средних широтах (около 3–4 мес) на поверхность Земли приходит около 2×109 ккал фотосинтетически активной радиации (ФАР; в области спектра от 380 до 720 нм). Из них в урожае биомассы в 10 т запасается около 40×106 ккал, т. е. 2% ФАР. Остальная энергия частично отражается, но в большей части превращается в тепло и вызывает испарение громадных количеств 2. Т. о., для усиления фотосинтетической деятельности растений необходимо повысить коэффициент использования растениями солнечной радиации. Это достигается увеличением в посевах размеров листовой поверхности, удлинением сроков активной деятельности листьев, регулированием густоты стояния растений. Важное значение имеет способ размещения растений на площади (правильные нормы высева семян), обеспечение их достаточным количеством 2 в воздухе, воды, элементов почвенного питания и т.д. Функциональная активность фотосинтетического аппарата, помимо внешних условий, определяется также анатомическим строением листа, активностью ферментных систем и типом метаболизма Большая роль принадлежит селекции растений – созданию сортов, обладающих высокой интенсивностью ассимиляции 2, и управлению процессами, связанными с эффективным использованием создаваемых при Ф органических веществ. Важное свойство высокопродуктивных сортов – способность использовать большую часть ассимилятов на формирование ценных в хозяйственном отношении органов (зерна у злаков, клубней у картофеля, корней у корнеплодов и т.д.). Выяснение законов и основ фотосинтетической продуктивности растений, разработка принципов ее оптимизации и повышения – важная задача современности.
Лит.: Любименко В. Н,, Ф и хемосинтез в растительном мире, М. – Л., 1935; Тимирязев К. А., Солнце, жизнь и М., 1937 (Соч., т. 1–2); Годнев Т. Н., Строение и возможные пути его образования в растении, М. – Л., 1947 (Тимирязевское чтение. 7); Теренин А. Н., Фотохимия и фотосинтез, М., 1951 (Баховское чтение. 6); Рабинович Е., Ф, пер. с англ., т. 1–3, М., 1951–59; Ничипорович А. А., Ф и теория получения высоких урожаев, М., 1956 (Тимирязевское чтение, 15); Воскресенская Н. П., Ф и спектральный состав света, М., 1965; Андреева Т. Ф, Ф и обмен листьев, М., 1969; Теоретические основы фотосинтетической продуктивности, Сб. докл. на Междунар. симпозиуме, М., 1972; Современные проблемы фотосинтеза. К 200-летию открытия фотосинтеза, М., 1973; Красновский А. А., Преобразование энергии света при фотосинтезе. Молекулярные механизмы, М., 1974 (Баховское чтение. 29); Фотохимические системы К., 1975; Bioenergetics of photosynthesis, . . – L. – Los Ang., 1975.
А. А. Ничипорович.
|
Для поиска, наберите искомое слово (или его часть) в поле поиска
|
|
 |
 |
 |
|
|
|
Новости 27.01.2025 07:18:33
|
|
|
|
|
|
|
|
|
|
|


