
|

|
Большая Советская Энциклопедия (цитаты)
|

|
|
|
|
Нервная система | Нервная система (далее Н) совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и обеспечивающая функционирование организма как единого целого в его постоянном взаимодействии с внешней средой. Н воспринимает внешние и внутренние раздражения, анализирует эту информацию, отбирает и перерабатывает ее и в соответствии с этим регулирует и координирует функции организма.
Н образована главным образом нервной тканью, основной элемент которой - нервная клетка с отростками, обладающая высокой возбудимостью и способностью к быстрому проведению возбуждения.
Эволюция Н, претерпеваемая ею в ходе филогенеза, отличается большой сложностью. У простейших - одноклеточных организмов - Н отсутствует, но у некоторых инфузорий имеется внутриклеточная сеточка, выполняющая функцию проведения возбуждения к др. элементам клетки. Самая примитивная форма Н, сохранившаяся лишь у низших кишечнополостных (гидра), - диффузная Н (рис. 1, А). Нервные клетки у кишечнополостных при помощи отростков соединяются в сеть, в которой проведение возбуждения может осуществляться во всех направлениях. В процессе дальнейшей эволюции строение Н усложняется. У свободно живущих кишечнополостных происходит образование и погружение в глубь тела скоплений нервных клеток - нервных узлов (ганглиев), - связи между которыми устанавливаются преимущественно при помощи длинных отростков (нервных волокон, нервов). Появление такого диффузно-узлового типа строения (рис. 1, Б) сопровождается развитием специализированных воспринимающих нервных структур (рецепторов), дифференцирующихся в соответствии с воспринимаемым ими видом энергии. Проведение возбуждения становится направленным. Следующий этап эволюции (кольчатые черви, членистоногие, иглокожие, моллюски) - переход к узловому типу строения Н: нервные клетки сосредоточены в узлах, связанных нервными волокнами между собой, а также с соответствующими рецепторами и исполнительными органами. Появляется дистантная рецепция; среди нервных узлов происходит выделение доминирующих, расположенных у свободно передвигающихся животных на головном конце тела и связанных с наиболее важными дистантными рецепторами - органами чувств (рис. 1, В). В связи с тем, что головные узлы получают при движении животного наибольшее количество информации от внешнего мира, они увеличиваются, структура их усложняется; туловищные ганглии приближаются к головным и сливаются с ними, образуя сложные мозговые комплексы, которые в какой-то мере подчиняют себе деятельность др. узлов.
У позвоночных животных тип строения Н резко отличается от узлового типа, обычно присущего беспозвоночным. Центральная нервная система (ЦНС) представлена нервной трубкой, расположенной на спинной стороне тела, и состоит из спинного и головного мозга. В эмбриональном развитии позвоночных Н образуется из наружного зародышевого листка - эктодермы (сперва в виде нервной пластинки, сворачивающейся в желобок, а затем превращающейся в нервную трубку) (рис. 2). Зачаточные эктодермальные клетки дифференцируются на нейробласты (клетки, дающие начало нейронам) и спонгиобласты (образующие клетки нейроглии). Из эктодермальных клеток, путем их миграции, формируются и периферические узлы, а совокупность отростков некоторых нейробластов образует черепномозговые и спинномозговые нервы, относимые к периферической Н Головной конец нервной трубки делится на 3 мозговых пузыря - зачатки головного мозга. В процессе развития передний мозговой пузырь разделяется на два, из которых один образует конечный мозг, включающий большие полушария и базальные ганглии, а второй - промежуточный мозг. Средний мозговой пузырь дает начало среднему мозгу. Из заднего - образуются мозжечок, варолиев мост и продолговатый мозг. Остальная часть нервной трубки, сохраняя трубчатое строение, образует спинной мозг с утолщениями в поясничной и плечевой областях. Как спинной, так и головной мозг позвоночных покрыт рядом оболочек и заключен в костные покровы - череп и позвоночник.
В процессе эволюции происходит дальнейшее усложнение структуры Н и усовершенствование всех форм ее взаимодействия с внешней средой; при этом все большее значение приобретают прогрессирующие в своем развитии передние отделы головного мозга. У рыб передний мозг почти не дифференцирован, но у них хорошо развиты задний, а также средний мозг; наибольшего развития у рыб достигает мозжечок. У земноводных и пресмыкающихся задний мозг занимает относительно меньший объем, чем у рыб, мозжечок же уступает в развитии среднему мозгу, который делится на 2 части, образуя двухолмие. Усложняется структурно и функционально передний мозговой пузырь, он дифференцируется на промежуточный мозг и 2 полушария с развитой нервной тканью, образующей так называемую первичную кору мозга. Передний мозг, первоначально связанный лишь с обонянием, затем приобретает и более сложные функции. Несколько обособленное место в эволюционном ряду занимают птицы, у которых доминируют структуры так называемого мозгового ствола, т. е. средний мозг и только те части переднего мозга, которые расположены в глубине полушарий (базальные ганглии, промежуточный мозг); сильно развит у птиц и мозжечок; кора головного мозга дифференцирована слабо.
Высшего развития Н достигает у млекопитающих. Головной конец нервной трубки в эмбриогенезе делится у них на 5 пузырей: передний - дает начало большим полушариям и промежуточному мозгу, средний - среднему мозгу, задний - делится на собственно задний (варолиев мост и мозжечок) и продолговатый мозг. Кора больших полушарий головного мозга образует многочисленные борозды и извилины. Первичная полость нервной трубки превращается в желудочки мозга и спинномозговой канал. Нейронная организация мозга крайне усложняется. Развитие и дифференциация структур Н у высокоорганизованных животных обусловили ее разделение на соматическую и вегетативную нервную систему. Особенность строения вегетативной Н та, что ее волокна, отходящие от ЦНС, не доходят непосредственно до эффектора, а сначала вступают в периферические ганглии, где оканчиваются на клетках, отдающих аксоны уже непосредственно на иннервируемый орган. В зависимости от того, где расположены ганглии вегетативной Н, и некоторых ее функциональных особенностей вегетативную Н делят на 2 части: парасимпатическую и симпатическую.
Структурная и функциональная единица Н - нейрон, состоящий из тела нервной клетки и отростков - аксона и дендритов. Кроме нервных клеток, в структуру Н входят глиальные клетки (см. Нейроглия). Нейроны являются в известной мере самостоятельными единицами - их протоплазма не переходит из одного нейрона в другой (см. Нейронная теория). Взаимодействие между нейронами осуществляется благодаря контактам между ними (см. Синапсы; рис. 3). В области контакта между окончанием одного нейрона и поверхностью другого в большинстве случаев сохраняется особое пространство - синаптическая щель - шириной в несколько сот 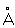 . Основные функции нейронов: восприятие раздражений, их переработка, передача этой информации и формирование ответной реакции. В зависимости от типа и хода нервных отростков (волокон), а также их функций нейроны подразделяют на: а) рецепторные (афферентные), волокна которых проводят нервные импульсы от рецепторов в ЦНС; тела их находятся в спинальных ганглиях или ганглиях черепномозговых нервов; б) двигательные (эфферентные), связывающие ЦНС с эффекторами; тела и дендриты их находятся в ЦНС, а аксоны выходят за ее пределы (за исключением эфферентных нейронов вегетативной Н, тела которых расположены в периферических ганглиях); в) вставочные (ассоциативные) нейроны, служащие связующими звеньями между афферентными и эфферентными нейронами; тела и отростки их расположены в ЦНС. . Основные функции нейронов: восприятие раздражений, их переработка, передача этой информации и формирование ответной реакции. В зависимости от типа и хода нервных отростков (волокон), а также их функций нейроны подразделяют на: а) рецепторные (афферентные), волокна которых проводят нервные импульсы от рецепторов в ЦНС; тела их находятся в спинальных ганглиях или ганглиях черепномозговых нервов; б) двигательные (эфферентные), связывающие ЦНС с эффекторами; тела и дендриты их находятся в ЦНС, а аксоны выходят за ее пределы (за исключением эфферентных нейронов вегетативной Н, тела которых расположены в периферических ганглиях); в) вставочные (ассоциативные) нейроны, служащие связующими звеньями между афферентными и эфферентными нейронами; тела и отростки их расположены в ЦНС.
Деятельность Н основывается на двух процессах: возбуждении и торможении. Возбуждение может быть распространяющимся (см. Импульс нервный) или местным - нераспространяющимся, стационарным (последнее открыто Н. Е. Введенским в 1901) Торможение - процесс, тесно связанный с возбуждением и внешне выражающийся в снижении возбудимости клеток. Одна из характерных черт тормозного процесса - отсутствие способности к активному распространению по нервным структурам (явление торможения в нервных центрах впервые было установлено И. М. Сеченовым в 1863).
Клеточные механизмы возбуждения и торможения подробно изучены. Тело и отростки нервной клетки покрыты мембраной, постоянно несущей на себе разность потенциалов (так называемый мембранный потенциал). Раздражение расположенных на периферии чувствительных окончаний афферентного нейрона преобразуется в изменение этой разности потенциалов (см. Биоэлектрические потенциалы). Возникающий вследствие этого нервный импульс распространяется по нервному волокну и достигает его пресинаптического окончания, где вызывает выделение в синаптическую щель высокоактивного вещества - медиатора. Под влиянием последнего в постсинаптической мембране, чувствительной к действию медиатора, происходит молекулярная реорганизация поверхности. В результате постсинаптическая мембрана начинает пропускать ионы и деполяризуется, вследствие чего на ней возникает электрическая реакция в виде местного возбуждающего постсинаптического потенциала (ВПСП), вновь генерирующего распространяющийся импульс. Нервные импульсы, возникающие при возбуждении особых тормозящих нейронов, вызывают гиперполяризацию постсинаптической мембраны и, соответственно, тормозящий постсинаптический потенциал (ТПСП). Помимо этого, установлен и др. вид торможения, формирующийся в пресинаптической структуре, - пресинаптическое торможение, обусловливающее длительное снижение эффективности синаптической передачи (см. Мембранная теория возбуждения).
В основе деятельности Н лежит рефлекс, т. е. реакция организма на раздражения рецепторов, осуществляемая при посредстве Н Термин "рефлекс" был впервые введен в зарождавшуюся физиологию Р. Декартом в 1649, хотя конкретных представлений о том, как осуществляется рефлекторная деятельность, в то время еще не было. Такие сведения были получены лишь значительно позже, когда морфологи приступили к исследованию строения и функций нервных клеток (Р. Дютроше, 1824; К. Эренберг, 1836; Я. Пуркине, 1837; К. Гольджи, 1873; С. Рамон-и-Кахаль, 1909), а физиологами были изучены основные свойства нервной ткани (Л. Гальвани, 1791; К. Маттеуччи, 1847; Э. Дюбуа-Реймон, 1848-49; Н. Е. Введенский, 1901; А. Ф. Самойлов, 1924; Д. С. Воронцов, 1924; и др.). В конце 19 и начале 20 вв. были созданы карты расположения нервных центров и нервных путей в мозге, а также получены сведения об основных рефлекторных процессах и о локализации функций в мозге, с тех пор постоянно пополняемые и расширяемые (И. М. Сеченов, 1863; Н. А. Миславский, 1885; В. М. Бехтерев, 1903; И. П. Павлов, 1903; Ч. Шеррингтон, 1906; А. А. Ухтомский, 1911; И. С. Бериташвили, 1930; Л. А. Орбели, 1932; Дж. Фултон, 1932; Э. Эдриан, 1932; П. К. Анохин, 1935; К. М. Быков, 1941; Х. Мэгоун, 1946; и др.). Все рефлекторные процессы связаны с распространением возбуждения по определенным нервным структурам - рефлекторным дугам. Основные элементы рефлекторной дуги: рецепторы, центростремительный (афферентный) нервный путь, внутрицентральные структуры различной сложности, центробежный (эфферентный) нервный путь и исполнительный орган (эффектор). Различные группы рецепторов возбуждаются раздражителями разной модальности (т. е. качественной специфичности) и воспринимают раздражения, исходящие как из внешней среды (экстерорецепторы - органы зрения, слуха, обоняния и др.), так и из внутренней среды организма (интерорецепторы, возбуждающиеся при механических, температурных и др. раздражениях внутренних органов, мышц и др.). Нервные сигналы, несущие в ЦНС информацию от рецепторов по нервным волокнам, лишены модальности и обычно передаются в виде серии однородных импульсов. Информация о различных характеристиках раздражений кодируется изменениями частоты импульсов, а также приуроченностью нервной импульсации к определенным волокнам (так называемое пространственно-временное кодирование).
Совокупность рецепторов данной области тела животного или человека, раздражение которых вызывает определенный тип рефлекторной реакции, называется рецептивным полем рефлекса. Такие поля могут накладываться друг на друга. Совокупность нервных образований, сосредоточенных в ЦНС и ответственных за осуществление данного рефлекторного акта, обозначают термином нервный центр. На отдельном нейроне в Н может сходиться огромное число окончаний волокон, несущих импульсы от др. нервных клеток. В каждый данный момент в результате сложной синаптической переработки этого потока импульсов обеспечивается дальнейшее проведение лишь одного, определенного сигнала - принцип конвергенции, лежащий в основе деятельности всех уровней Н ("принцип конечного общего пути" Шеррингтона, получивший развитие в трудах Ухтомского и др.).
Пространственно-временная суммация синаптических процессов служит основой для различных форм избирательного функционального объединения нервных клеток, лежащего в основе анализа поступающей в Н информации и выработки затем команд для выполнения различных ответных реакций организма. Такие команды, как и афферентные сигналы, передаются от одной клетки к другой и от ЦНС к исполнительным органам в виде последовательностей нервных импульсов, возникающих в клетке в том случае, когда суммирующиеся возбуждающие и тормозящие синаптические процессы достигают определенного (критического для данной клетки) уровня - порога возбуждения.
Несмотря на наследственно закрепленный характер связей в основных рефлекторных дугах, характер рефлекторной реакции может в значительной степени изменяться в зависимости от состояния центральных образований, через которые они осуществляются. Так, резкое повышение или понижение возбудимости центральных структур рефлекторной дуги может не только количественно изменить реакцию, но и привести к определенным качественным изменениям в характере рефлекса. Примером такого изменения может служить явление доминанты.
Важное значение для нормального протекания рефлекторной деятельности имеет механизм так называемой обратной афферентации - информации о результате выполнения данной рефлекторной реакции, поступающей по афферентным путям от исполнительных органов. На основании этих сведений в случае, если результат неудовлетворителен, в сформировавшейся функциональной системе могут происходить перестройки деятельности отдельных элементов до тех пор, пока результат не станет соответствовать уровню, необходимому для организма (П. К. Анохин, 1935).
Всю совокупность рефлекторных реакций организма делят на две основные группы: безусловные рефлексы - врожденные, осуществляемые по наследственно закрепленным нервным путям, и условные рефлексы, приобретенные в течение индивидуальной жизни организма путем образования в ЦНС временных связей. Способность образования таких связей присуща лишь высшему для данного вида животных отделу Н (для млекопитающих и человека - это кора головного мозга). Образование условнорефлекторных связей позволяет организму наиболее совершенно и тонко приспосабливаться к постоянно изменяющимся условиям существования. Условные рефлексы были открыты и изучены И. П. Павловым в конце 19 - начале 20 вв. Исследование условнорефлекторной деятельности животных и человека привело его к созданию учения о высшей нервной деятельности (ВНД) и анализаторах. Каждый анализатор состоит из воспринимающей части - рецептора, проводящих путей и анализирующих структур ЦНС, обязательно включающих ее высший отдел. Кора головного мозга у высших животных - совокупность корковых концов анализаторов; она осуществляет высшие формы анализаторной и интегративной деятельности, обеспечивая совершеннейшие и тончайшие формы взаимодействия организма с внешней средой.
Н обладает способностью не только немедленно перерабатывать поступающую в нее информацию при помощи механизма взаимодействующих синаптических процессов, но и хранить следы прошлой активности (механизмы памяти). Клеточные механизмы сохранения в высших отделах Н длительных следов нервных процессов, лежащие в основе памяти, интенсивно изучаются.
Наряду с перечисленными выше функциями Н осуществляет также регулирующие влияния на обменные процессы в тканях - адаптационно-трофическую функцию (И. П. Павлов, Л. А. Орбели, А. В. Тонких и др.). При перерезке или повреждении нервных волокон свойства иннервируемых ими клеток изменяются (это касается как физико- свойств поверхностной мембраны, так и биохимических процессов в протоплазме), что, в свою очередь, сопровождается глубокими нарушениями в состоянии органов и тканей (например, трофическими язвами). Если иннервация восстанавливается (в связи с регенерацией нервных волокон), то указанные нарушения могут исчезнуть.
Изучением строения, функций и развития нервной системы у человека занимается неврология. Нервные болезни - предмет невропатологии и нейрохирургии.
Лит.: Орбели Л. А., Лекции по физиологии нервной системы, 3 изд., М. - Л., 1938; его же, Избр. труды, т. 1-5, М. - Л., 1961-68; Ухтомский А. А., Собр. соч., т. 1-6, Л., 1945-62; Павлов И. П., Полн. собр. соч., 2 изд., т. 2, М., 1951; Сеченов И. М., Избр. произв., т. 1, (М.), 1952; Коштоянц Х. С., Основы сравни тельной физиологии, т. 2, М., 1957; Бериташвили И. С., Общая физиология мышечной и нервной системы, 3 изд., т. 1, М., 1959; Сепп Е. К., История развития нервной системы позвоночных, 2 изд., М., 1959: Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; Беклемишев В. Н., Основы сравнительной анатомии беспозвоночных, 3 изд., т. 2, М., 1964; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Окс С., Основы нейрофизиологии, пер. с англ., М., 1969; Шеррингтон Ч., Интегративная деятельность нервной системы, пер. с англ., Л., 1969: Костюк П. Г., Физиология центральной нервной системы, К., 1971; Ariens Kappers С. ., Huber G. С., Crosby E. С., The comparative anatomy of the nervous system of vertebrates, including man, v. 1-2, . ., 1936; Bullock T. Н., Horridge G. A., Structure and function in the nervous systems of invertebrates, v. 1-2, . . - L., 1965.
П. Г. Костюк.
|
Для поиска, наберите искомое слово (или его часть) в поле поиска
|
|
 |
 |
 |
|
|
|
Новости 13.06.2026 16:20:15
|
|
|
|
|
|
|
|
|
|
|


